背景
在深層皮膚損傷后,哺乳動(dòng)物的進(jìn)化為了加速傷口愈合,會(huì)產(chǎn)生纖維化疤痕。我們無法再生功能性真皮以及一些功能附屬物是人類皮膚傷口愈合的主要障礙。有趣的是,一些組織擁有再生的能力,例如全層切除皮膚傷口后的新生毛囊(HF) 形成已經(jīng)在小鼠中被證明,這作為一個(gè)強(qiáng)大的模型,以了解潛在的皮膚再生機(jī)制。哺乳動(dòng)物的新皮膚修復(fù)纖維化瘢痕的形成是由于真皮成纖維細(xì)胞占優(yōu)勢(shì),容易產(chǎn)生過量的纖維胞外基質(zhì)。因此,確定成纖維細(xì)胞的來源對(duì)于改善傷口愈合效果的治療至關(guān)重要。
長期以來,HF 被認(rèn)為是傷口愈合過程中再生間充質(zhì)細(xì)胞的潛在來源。事實(shí)上,成人 HFs 內(nèi)真皮干細(xì)胞(hfDSCs),在 HFs 經(jīng)歷反復(fù)的變性、重塑和再生周期時(shí),能夠不斷地用新的間充質(zhì)干細(xì)胞(mesenchymal cells)重新填充 HFs。在這里,作者想知道 HF 相關(guān)的間充質(zhì)祖細(xì)胞(mesenchymal progenitors,MPs)是否參與了傷口愈合,以及它們是否代表了新生 HFs 中的誘導(dǎo)間充質(zhì)干細(xì)胞的主要來源。已有研究發(fā)現(xiàn),骨骼肌和心臟中存在 MPs,Hic1 是 MPs 的 maker 基因,在組織損傷后被激活。
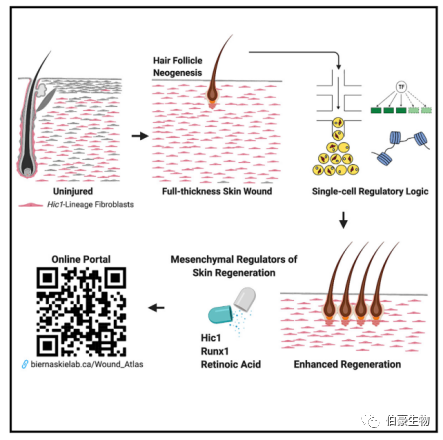
摘要
真皮成纖維細(xì)胞在穩(wěn)態(tài)時(shí)和損傷時(shí)的反應(yīng)表現(xiàn)出相當(dāng)大的異質(zhì)性。定義修復(fù)性成纖維細(xì)胞的譜系起源和促進(jìn)纖維化或相反地促進(jìn)毛囊再生的調(diào)節(jié)程序?qū)?duì)促進(jìn)傷口愈合效果至關(guān)重要。作者發(fā)現(xiàn)毛囊間 MPs 對(duì)創(chuàng)傷修復(fù)的作用有限。相反,以 Hic1 為標(biāo)志的毛囊間的祖細(xì)胞產(chǎn)生了大量的修復(fù)性成纖維細(xì)胞,并表現(xiàn)出功能分化,介導(dǎo)創(chuàng)面新真皮中心的再生和周圍瘢痕的形成。通過 scRNA-seq 發(fā)現(xiàn)了獨(dú)特的轉(zhuǎn)錄、調(diào)控和上皮 - 間充質(zhì)轉(zhuǎn)化特征使 MPs 具有再生能力。同時(shí)結(jié)合 scATAC-seq 突出了再生相關(guān)基因座內(nèi)染色質(zhì)可及性的變化。針對(duì) RUNX1 和維甲酸信號(hào)的藥理調(diào)節(jié)或在創(chuàng)傷激活的成纖維細(xì)胞中,同時(shí)發(fā)現(xiàn) Hic1 的缺失促進(jìn)愈合,表明修復(fù)性成纖維細(xì)胞具有潛在可變化的再生能力。
實(shí)驗(yàn)設(shè)計(jì)
scRNA-seq
為了分離包含創(chuàng)面新真皮的細(xì)胞,作者在 Hic1-tdTomato+ 小鼠幼鼠身上創(chuàng)建了小(8 毫米直徑)和大(1.5 厘米直徑)的全切層傷口。通過流式分選獲得 5 個(gè)關(guān)注的細(xì)胞群(1. 小創(chuàng)面 D8 (SWD8) 細(xì)胞,2.SWD14, 3.D14 大創(chuàng)面外周細(xì)胞,4.D14 大創(chuàng)面中心細(xì)胞,5.P28 時(shí)期未傷皮膚細(xì)胞),即 5 個(gè)樣本,然后分別上機(jī) 10X Genomics 進(jìn)行 scRNA-seq。
scATAC-seq
分離細(xì)胞核用于 10x Chromium Next GEM scATAC -seq 分析,從 D14 大傷口中心區(qū)域中收集了 50,000 個(gè)細(xì)胞,放入 100ml 的冷凍裂解緩沖液中,在冰上抽核,要求無核起泡。使用 CountessTM II 自動(dòng)細(xì)胞計(jì)數(shù)儀對(duì)細(xì)胞核進(jìn)行定量計(jì)數(shù),使用 10x Chromium Chip E 對(duì) 5000 個(gè)細(xì)胞核進(jìn)行捕獲,并進(jìn)行后續(xù)的測(cè)序。
結(jié)果展示
1. 在一定的微環(huán)境中,毛囊間成纖維細(xì)胞 HE Hic1-Lineage-Comprising HFs 有助于 HFs 再生
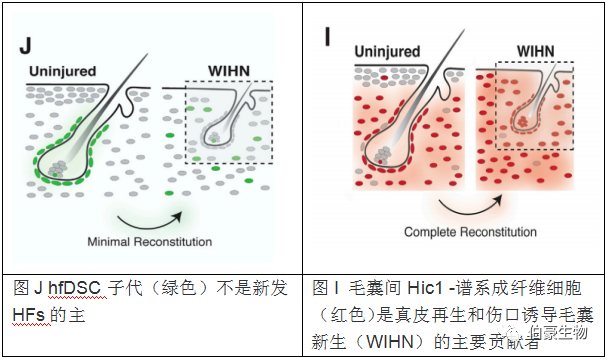
2. 單細(xì)胞轉(zhuǎn)錄組學(xué)顯示不同的分子程序調(diào)節(jié)成纖維細(xì)胞對(duì)損傷的反應(yīng)
為了剖析在纖維化和再生愈合過程中驅(qū)動(dòng)成纖維細(xì)胞分化的分子程序,我們從未受傷的皮膚(P28)、8 和 14dpw 的小傷口和 14dpw 的大傷口(分為中央和外周區(qū)域)中分析了 29,269 個(gè)單細(xì)胞。
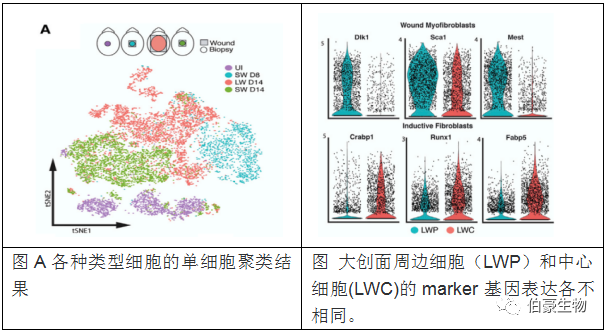
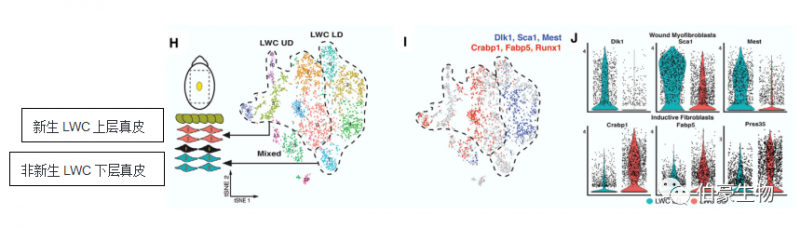
作者對(duì)大面積創(chuàng)傷對(duì)的中心區(qū)域細(xì)胞單獨(dú)聚類得到新生 LWC 上層真皮和非新生 LWC 下層真皮。Crabp1+/Prss35+ 成纖維細(xì)胞定位于新生 LWC 上層真皮。
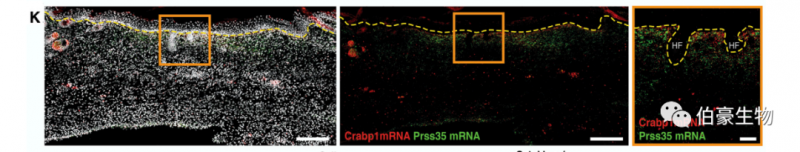
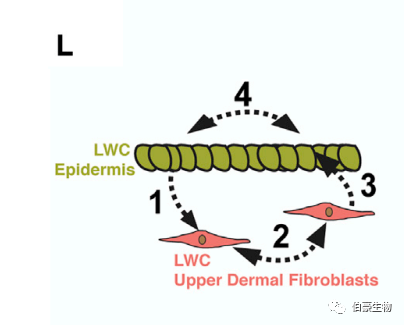
這表明,成纖維細(xì)胞的空間分離及其與上覆表皮基底細(xì)胞的相互作用是獲得再生能力所必需的。由于活性成纖維細(xì)胞和表皮干細(xì)胞之間的上皮 - 間充質(zhì)細(xì)胞的相互作用可啟動(dòng)胚胎皮膚中新生 HF 的形成,作者推斷類似的調(diào)控相互作用可能建立再生能力,使 WIHN 成為可能。通過使用 CellPhoneDB 將 14 DPW 的 LWC 上層真皮成纖維細(xì)胞與 LWC 表皮細(xì)胞進(jìn)行分析發(fā)現(xiàn)了一個(gè)獨(dú)特的相互作用體,讓人聯(lián)想到 HF 形態(tài)形成過程中的信號(hào)交換。例如,ephrin 受體(即 Efna2、Efna4 和 Efnb1) 和配體(即 Epha1、Epha4、Epha5、Ephb2 和 Ephb6) 被認(rèn)為是胚胎皮膚形態(tài)形成過程中表皮增殖的負(fù)調(diào)節(jié)因子(Genander, Holmberg 和 Frise’n, 2010),預(yù)測(cè)將在再生的新真皮內(nèi)相互作用,表明保守調(diào)控相互作用的再激活使間充質(zhì)具有再生能力(圖 L)。
3. 在新生毛囊域中表觀遺傳譜特征可區(qū)分成纖維細(xì)胞狀態(tài)
表觀基因組可能是再生愈合過程中成纖維細(xì)胞功能的關(guān)鍵決定因素,這意味著毛囊間成纖維細(xì)胞在早期具有通過表觀基因組的改變以重新激活真皮轉(zhuǎn)錄狀態(tài)并獲得誘導(dǎo)間充質(zhì)干細(xì)胞對(duì)功能的潛在能力雖然轉(zhuǎn)錄組的改變可能是皮膚和 HF 再生的必要條件,但不足以激活關(guān)鍵 TFs 的下游靶點(diǎn)。因此作者認(rèn)為成纖維細(xì)胞可能通過調(diào)節(jié)染色質(zhì)狀態(tài)可及性來激活或阻斷 TF 活性以調(diào)節(jié)下游的基因表達(dá)。為了直接測(cè)試這兩種可能性,作者使用 scATAC-seq 檢測(cè) LWC 14 dpw 樣本的染色質(zhì)可及性。
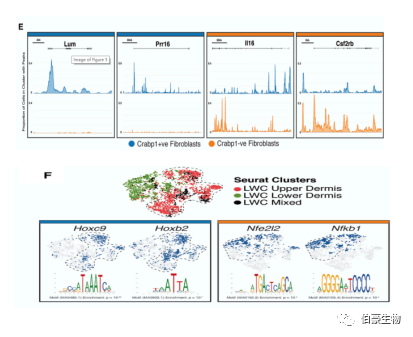
上層成纖維細(xì)胞可表達(dá) Pdgfra 和 Crabp1 和下層成纖維細(xì)胞可表達(dá) Pdgfra 但不能表達(dá) Crabp1。有趣的是,作者發(fā)現(xiàn)胚胎活性蛋白多糖 Lum 和代謝調(diào)節(jié)因子 Prr16 在啟動(dòng)子區(qū)對(duì)的染色質(zhì)開放是可表達(dá) Pdgfra 和 Crabp1 的上層成纖維細(xì)胞染色質(zhì)特征的一部分(圖 E)。在 579 個(gè)差異富集 motif 中,作者發(fā)現(xiàn)許多 TFs 被同時(shí)在上、下成纖維細(xì)胞中預(yù)測(cè)為具有差異可及性和活性(圖 F)。
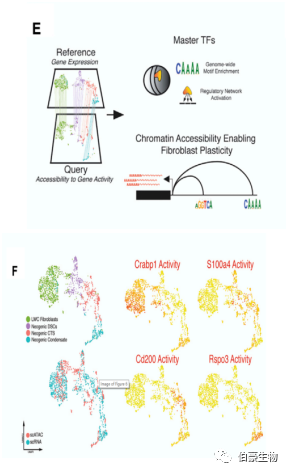
緊接著作者希望找出 scATAC -seq 和 scRNA-seq 之間的對(duì)應(yīng)關(guān)系是否能夠揭示毛囊相關(guān)的表觀遺傳特征。作者將 LWC 14 dpw 上層真皮細(xì)胞的 scATAC-seq 與 scRNA-seq 進(jìn)行整合分析。作者的確發(fā)現(xiàn)新生的 HF 間充質(zhì)干細(xì)胞的狀態(tài)有著不同的染色質(zhì)譜,有助于實(shí)現(xiàn)下游精準(zhǔn)的轉(zhuǎn)錄調(diào)控(圖 E 和 F)。
例如,Grem2 在 LWC 成纖維細(xì)胞的整個(gè)基因體上高度可及(圖 F),但伴隨 HF 間充質(zhì)細(xì)胞的發(fā)生后,其染色質(zhì)可及性丟失。總之,作者的多組學(xué)特性揭示了調(diào)控成纖維細(xì)胞的上游機(jī)制,并列出了其可能作為成功刺激真皮再生和 / 或減輕纖維化的重要分子靶點(diǎn)的關(guān)鍵 TFs。
4. 單細(xì)胞多組學(xué)顯示 Runx1 和維甲酸是間充質(zhì)細(xì)胞再生的主要調(diào)節(jié)因子
為了評(píng)估通過多組學(xué)分析鑒定的調(diào)控因子的功能意義,作者將維甲酸(RA) 和 Runx1 途徑列入了候選名單,因?yàn)轵?qū)動(dòng)這些通路的幾個(gè) TFs 只在再生 LWC 上層真皮細(xì)胞中活躍(Figures BCD)。
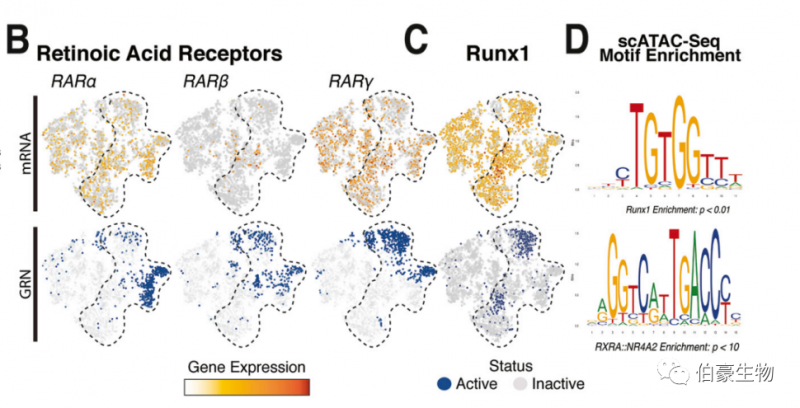

為了確定 RA 和 Runx1 信號(hào)通路是能激活 WIHN,我們將小分子抑制劑用于傷口愈合實(shí)驗(yàn)。圖 F 顯示 RA 和 Runx1 抑制劑減少了再生毛囊數(shù)量。相反,外源性 RA 的應(yīng)用增加了再生能力,導(dǎo)致新生 HFs 增加 1.5 倍。
為了使確定的候選基因能夠廣泛傳播,作者還建立了一個(gè)可搜索的網(wǎng)站,名為 Wound Atlas((http://www.biernaskielab.ca/wound_atlas)。隨著更多數(shù)據(jù)的積累,這個(gè)平臺(tái)將會(huì)定期更新,作者希望能激發(fā)對(duì)跨組織間充質(zhì)動(dòng)力學(xué)基因調(diào)節(jié)的研究。
5. Hic1 缺失通過增加 LWs 再生區(qū)域的成纖維細(xì)胞密度來增強(qiáng) WIHN
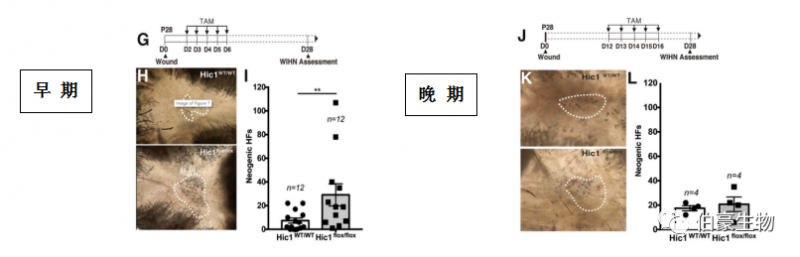
因?yàn)?Hic1 可調(diào)節(jié) MP 的狀態(tài),其失活導(dǎo)致骨骼肌和心臟損傷后 MP 過度活化,從而導(dǎo)致纖維化,作者想知道 Hic1 是否同樣會(huì)改變皮膚創(chuàng)面的愈合結(jié)果。為了驗(yàn)證,作者構(gòu)建了 Hic1 缺失的模型。有趣的是,雖然在創(chuàng)傷早期(D2-6)Hic1 失活導(dǎo)致新生 HFs 數(shù)量急劇增加 4 倍(圖 GHI)、晚期失活(D12-16) 對(duì) WIHN 無影響(圖 7K 7M)。
展望
上述研究這支持了這樣一種觀點(diǎn),即 Hic1 缺陷本身并不會(huì)使成纖維細(xì)胞產(chǎn)生親纖維化反應(yīng),相反,當(dāng)暴露在適當(dāng)?shù)脑试S環(huán)境中,成纖維細(xì)胞卻能夠適應(yīng)再生能力。在傷口愈合過程中,短暫調(diào)節(jié) Hic1 以更有效地調(diào)動(dòng)皮膚 MPs,并提供一個(gè)寬松(即促進(jìn)促進(jìn)毛囊再生)的傷口環(huán)境,其可能是一種提高傷口愈合效果的可行治療方法。
(原文鏈接:https://doi.org/10.1016/j.stem.2020.07.008)
文章不足
作者將控制性別(female)的生物學(xué)重復(fù)創(chuàng)傷組織用于 scRNA-seq,這樣就排除了對(duì)纖維母細(xì)胞狀態(tài)的生物學(xué)或性別特異性差異的公正評(píng)估。盡管來自 LWC 細(xì)胞的 scATAC-seq 比較了再生能力與非再生成纖維細(xì)胞,但這樣排除了在招募到不同創(chuàng)傷區(qū)域(LWC 與 LWP) 或創(chuàng)傷類型(SWs 與 LWs) 的成纖維細(xì)胞間差異染色質(zhì)狀態(tài)可及性 /motiif 的分析。
伯豪生物為您提供專業(yè)的 10X genomics 平臺(tái)的 scATAC-seq 測(cè)序服務(wù),更能提供全面的 scRNA-seq 和 scATAC-seq 的聯(lián)合分析。詳情請(qǐng)咨詢 Market@shbio.com。
更多伯豪生物人工服務(wù):

